一般情況下天擇偏好子代數量較多的個體,其子代也能產生較多後代,因此生產力高的性狀便被保留下來。然而因環境與物種特性的交互下,生物在生活史個階段承受不同天擇壓力,其他生殖策略亦可能有相同的適應性而被天擇保留。生物的生殖投資策略,可藉由能量與時間分配的權衡 (trade-off) 解釋不同策略的演化優勢。在總能量相同的條件下,增加每個卵平均分配到的能量可增加存活率,但減少子代數量;提高總子代數量則降低子代存活率。前者除了將能量用在產生較大的卵之外,許多生物藉由額外的行為改變,提高子代存活率。包括交配前的求偶或築巢行為,以及交配後的護衛與餵食等。交配後的生殖投資稱為親代撫育 (parental care),包括胎生、護衛、搬運、餵食等。親代撫育必須犧牲額外的交配機會及自身營養,例如雌眼鏡王蛇在護卵期間無法覓食,會減少大量體重。有些物種可能因妊娠而增加額外的體重或活動頻率,而提高暴露於掠食者攻擊的危險。在子代存活率與多方風險的適應性衝突下,理論上天則會偏好適應性最佳的策略。因此親代撫育演化出高度多樣性,源於不同生物會表現出各自的親代撫育行為,並隨環境變動呈現不同的分佈。
兩棲類生物具有高度多樣的親代撫育行為,表現在行為複雜度、撫育時間、撫育性別。多數演化支系之內的物種,表現之撫育模式大多相似。如哺乳類多為胎生且撫育時間長 (相對於生活史);鳥類多為雙親共育;魚類則多為雄性撫育。然而對兩棲類親代撫育則不存在類似的分佈模式,各種撫育行為散佈在各演化支系,似乎是多次獨立演化事件,且同類型的撫育行為在不同支系間存在細微差異。例如孵卵行為便存在口孵、胃孵、背孵等多種模式,其相應生理或形態適應便有不同的特化。背孵卵的產婆蟾 (Alytes spp.) 雄性在育幼期間蟄伏於潮濕陰暗處直到卵孵化為蝌蚪。胃孵卵的胃育溪蛙 (Rheobatrachus silus) 在育幼期間消化機能會暫停,且可觀察到腺體退化,直到卵孵育至亞成體。兩者撫育行為除了生殖投資的程度不同,其複雜度與牽涉的生理變化亦有很大差異,可預期會對天擇變化衍伸出不同的反應,而有不同演化速率與可逆性。產婆蟾主要為行為適應,可能牽涉到較少的適應性突變。且棲地的幼體捕食者較少,卵對親代的依賴性也較低,因此該撫育行為可能在天擇壓力降低時消失。胃育溪蛙相對牽涉複雜的生理調控,並有明顯的形態改變,應為較多基因控制。且幼體對此撫育行為的依賴性較高,若此行為消失,則將造成適應性下降,因此無論外部天擇壓力是否減少,可逆性仍較低。儘管學界有大量對各種兩棲類親代撫育的研究與描述,目前對撫育行為在整個兩棲類支系的演化歷程了解不多。
Furness et al. 2019 在兩棲類演化樹重建各種親代撫育行為在各支系上的分佈,並藉由演化模型分析各種特徵的演化速率,並以此解釋各行為模式適應上的穩定性及彼此的交互關係。然而複雜且多樣的特徵不容易編碼 (coding),若過於粗略的歸類撫育行為,可能無法完整呈現演化歷程;但詳細歸類則會因雜訊過多難以歸納演化趨勢;而錯誤的歸類則會做出偏離事實的推論。Furness 的團隊將一千三百多種兩棲類的親代撫育行為大致分為護衛、搬運、孵育、餵食、胎生。護衛行為主要為在旁看守並驅趕掠食者、寄生者,有些會維持清潔與濕潤避免真菌感染或脫水。例如玻璃蛙科的 Hyalinobatrachium spp. 雄蛙會用身體遮蓋卵,避免掠食者或寄生者危害。然而本研究所回顧的樹蛙屬 (Rhacophorus) 之十九個物種皆會製造卵泡保護卵,且產卵後會短暫停留在卵附近,但因無明顯保護行為而編碼為無親代撫育。搬運行為大多在變動較大的環境,將幼體搬移至適合生長處。如 Dentrobates pumilio 因為卵與蝌蚪發育的環境不同,因此雌蛙會等卵孵化後在搬運到鳳梨科植物的積水處。孵育包括將幼體或卵負於背上或直養在口腔或鳴囊中,隔絕外界危險,並在必要時進行搬運。如前文所述的胃育溪蛙等。餵食多為雌性產生未受精的卵,提供為蝌蚪或亞成體的食物,常見於毒蛙科 (Dendrobatidae) 物種。但亦有部分物種由雄性或兩性提供食物。另外卵生的蚓螈 Boulengerula spp. 則由雌性提供增生的表皮作為亞成體的食物。胎生為幼體在母體子宮內發育至蝌蚪或亞成體,一般在描述涉及複雜營養交換系統的生殖模式,因此胃孵或背孵皆不屬於胎生。多數蚓螈及部分蠑螈,以及少數蟾蜍科 (Bufonidae) Nectophrynoides 屬之物種為胎生。
作者同時將被撫育之生活史階段納入分析,全部分為十種撫育行為:卵護衛 (EA)、蝌蚪護衛 (TA)、亞成體護衛 (JA)、蝌蚪搬運(TT)、亞成體搬運 (JT)、卵孵育 (EB)、蝌蚪孵育 (TB)、蝌蚪餵食 (TF)、亞成體餵食 (JF)、胎生 (V)。該研究所包含的物種皆無卵搬運 (ET) 之紀錄,因此未被涵蓋。亞成孵育 (JB) 之行為大多被包含在 JT 之中,且有時為 EB 或 TB 之延伸,因此不獨立為一類做分析。然而許多撫育行為有部份重疊,此研究雖對撫育行為做詳盡的回顧,但要將生物的行為模式歸類成人為定義的框架,仍有一定的困難及限制。例如南美洲的負子蟾 (Pipa spp.) 直接將卵包埋於雌蟾背部皮膚,直到發育至亞成體,便包含了孵育、護衛、餵食的行為元素。然而作者的編碼將這種模式列為卵孵育,以方便資料處理,日後研究不能排除忽視重要演化資訊的可能。
研究發現不同類別之親代撫育行為在演化樹上的產生與消失速率有各自的模式,大致的趨勢為護衛行為產生與消失之速率最快;餵食與搬運中等;胎生與孵育最慢 (Fig. 1)。支持模式較簡單的撫育行為,因牽涉到的適應性突變較少,因此容易產生或消失。而伴隨複雜形態或生理適應的親代撫育,理論上牽涉到較多基因,需要較長的時間累積適應性突變因此產生速率慢,且根據適應性景觀 (fitness landscape) 理論,漸進的演化機制下不利於表徵逆向 (適應性較低的表徵) 演化,因此在支系中消失的速率也慢。例如胎生未在任何演化支系消失。

Fig. 1. 不同類別之撫育行為之演化速率分佈。a. 各行為在演化樹上之分布模式。b. 各行為產生速率。c. 各行為消失速率。( Furness et al. 2019)
若分析不同撫育行為針對的幼體階段,行為產生的速率也符合前述,但消失的速率卻較為多樣。此現象可能與不同階段的撫育行為之生殖投資權衡有關 (Fig. 2)。卵護衛行為產生快速,可能因為多數兩棲類卵期所受的天擇壓力大,而有較高的死亡率,因此卵護衛行為能有效提高適應性。然而又因為其所耗費的能量與時間較少,即使處於弱天擇壓力,卵護衛行為消失也不減少存活率的情況下,也不會因省下的時間與能量提高適應性,因此消失的速率相較於對蝌蚪期的護衛行為慢。蝌蚪期與亞成體期,因為不像卵固著不動,能主動逃離危險,可預期死亡率比卵低,因此撫育行為對存活率的提升有限。另一方面蝌蚪因為能量不再由卵黃提供,親代可能需要負擔餵食與或搬運等撫育行為,需要更高的生殖投資,在弱天擇壓力下可能消失。雖然蝌蚪期的撫育行為大多是卵期撫時間延伸,但對適應性提升的邊際效應遞減,因此在未牽涉形態適應之較為簡單的護衛及餵食行為,產生速率比卵期撫育慢,消失速率卻較快。較為複雜的孵育行為在卵期與蝌蚪期消失速率則相近。
搬運行為仰賴親代的空間記憶能力,比起留在原地的護衛行為較為複雜,產生速率較慢。另一方面,搬運行為存在親子適應性衝突以及子代間競爭。因為多數物種無法同時搬運所有子代,所以在兩趟間的空窗期,被搬運的個體適應性提高,將降低未被搬運之個體的適應性,同時也降低親代的總適應性,而存在一定的天擇敏感性。因此雖數複雜行為,但適應性的方向不同,消失速率中等 (介在胎生與護衛之間)。且無尾目的搬運行為主要集中在毒蛙類群 (Aromobatidae and Dendrobatidae),且大多處在天擇壓力大的雨林環境,因此在多數物種皆會呈現相似的演化速率。至於有尾目只有鰻螈科 (Sirenidae) 以及隱鰓鯢科 (Cryptobranchidae) 的日本大鯢兩個多樣性較低的支系表現亞成體搬運行為,支系間所呈現的演化速率也較為一致。
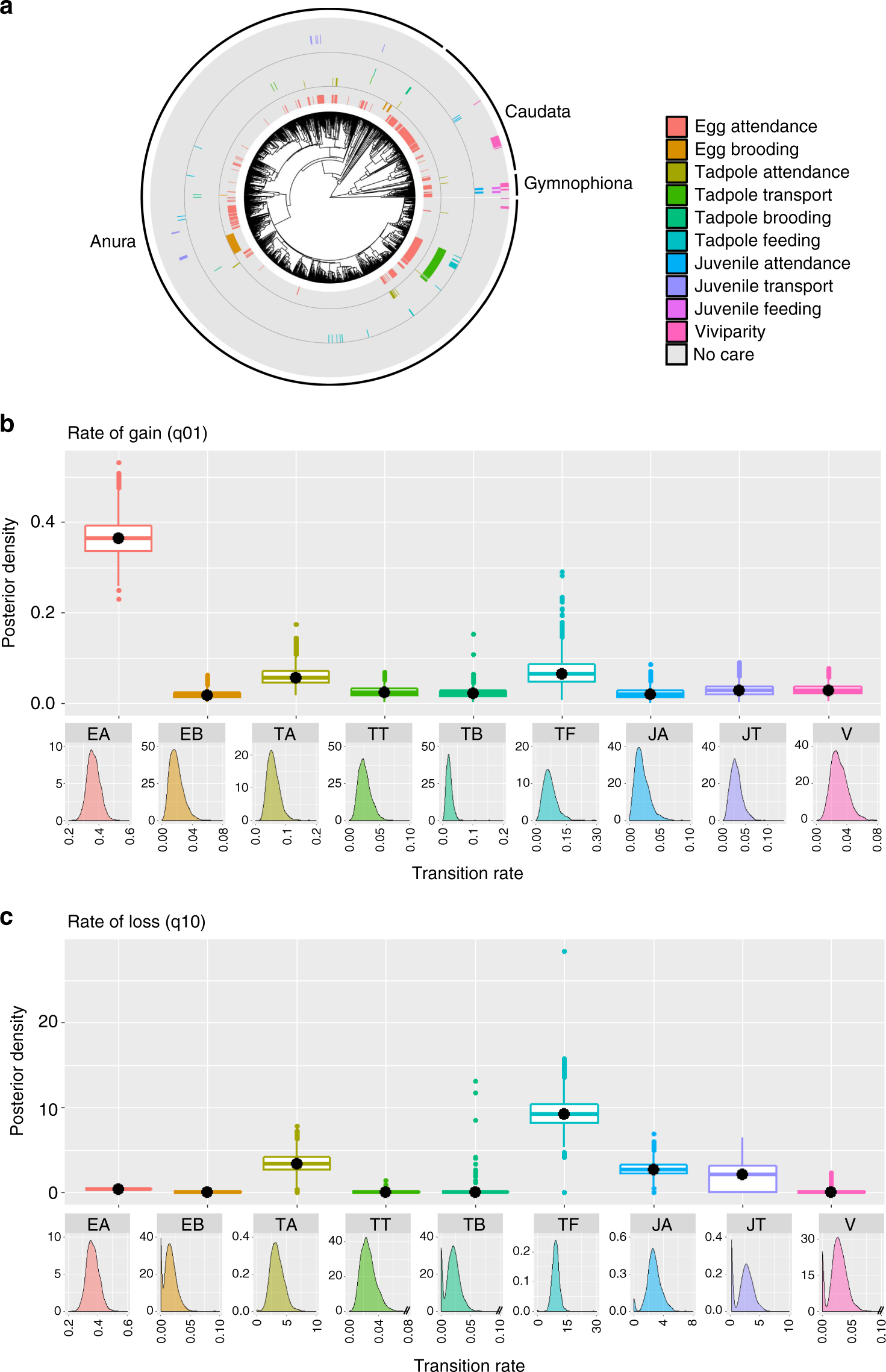
Fig. 2. 各生活史階段之不同類別之撫育行為之演化速率分佈。a. 各行為在演化樹上之分布模式。b. 各行為產生速率。c. 各行為消失速率。卵護衛 (EA)、蝌蚪護衛 (TA)、亞成體護衛 (JA)、蝌蚪搬運(TT)、亞成體搬運 (JT)、卵孵育 (EB)、蝌蚪孵育 (TB)、蝌蚪餵食 (TF)、亞成體餵食 (JF)、胎生 (V)。( Furness et al. 2019)
若只考慮被撫育之生活史階段轉變之演化速率如 Fig. 3 所示,只對生活史晚期撫育 (L) 的行為最不穩定,有高機率衍伸為全幼期撫育 (B) 或完全不撫育 (N)。不撫育、生活史前期撫育 (E) 以及全幼期撫育都不易消失或演化為其他模式。只對生活史晚期撫育大多發生在卵與蝌蚪與亞成體發育環境不同,如毒蛙科的蝌蚪在鳳梨科植物的積水中發育,因食物匱乏而面對較卵期更高的天則壓力,需要額外生殖投資,此行為模式容易衍伸為對卵期的護衛行為 (LB)。 然而對蝌蚪期的生殖投資較卵期高,因此在天擇壓力弱之情況下,很有可能促使此行為消失 (LN)。可推測撫育行為可能存在由死亡率最高的階段,拓展至死亡率較低的階段之趨勢。而不是單純透過卵期撫育延長時間之機制產生全幼期撫育。
雖然卵期撫育只需延長時間就會轉變為全幼期撫育,但如前文所述,需考量對適應性的權衡,因此生活史前期撫育演化為全幼期撫育的機率不高。只有在親子衝突與子代間競爭強的情況下,較容易由生活史前期撫育演化為全幼期撫育 (EB)。可能的機制為卵的費洛蒙對親代行為之影響或控制,讓親代在某些個體投入更多能量;或者在在子代數超過親代能搬運或餵食的數量,會產生較強的子代間競爭,具有較能操控親代的乞食行為可能促成撫育時間延長。可推論親代對資源分配的主動性越高,則撫育行為較容易因天擇壓力減弱而消失,如護衛行為。而可能牽涉到子代對親代行為操控,並影響適應性者如餵食、孵育、胎生,因為子代對撫育行為依賴提高,撫育行為的消失可能造成適應性降至零而較不可逆,為適應性景觀之一種呈現。
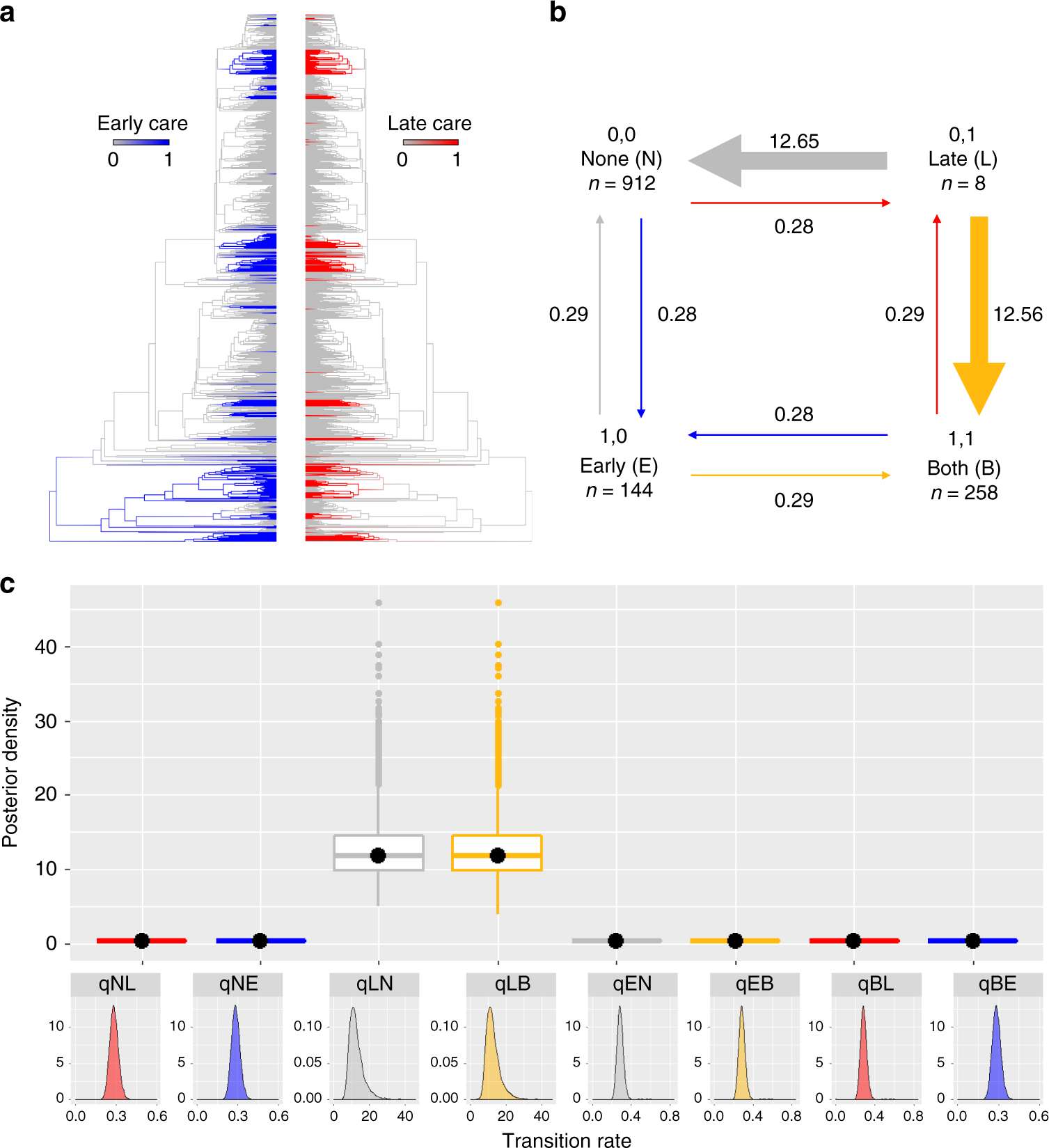
Fig. 3. 被撫育之生活史階段演化分佈。a. 各撫育時期於演化樹上之分布模式。b, c. 撫育行為執行時期之轉變機率。生活史前期撫育 (E)。活史晚期撫育 (L)。全幼期撫育 (B)。不撫育 (N)。( Furness et al. 2019)
雙親共育牽涉到兩性衝突與撫育分工,理論上若兩性撫育對子代個別之存活率相同,其中一方只要減少對同一巢的生殖投資,就會增加額外的適應性,例如額外的交配機會或降低撫育時面對的危險,提高下一次繁殖季適應性 (Smith 1977)。因此多數情況下,雙親共育由於存在適應性衝突,屬於演化不穩定的行為模式,應具有更高的消失速率。本研究模擬的結果高度支持此假說,雙親共育行為有高機率演化為單性撫育 (Fig. 4)。其中護衛與搬運行為,兩性對幼體的存活率之幫助相近,此趨勢更為明顯。然而,理論上兩性的撫育行為對存活率影響不同,或是工作互補的情況下,兩性共育消失速率應較慢,但資料未呈現此現象。可能只在極高的天擇壓力之下,才有可能穩定存在。
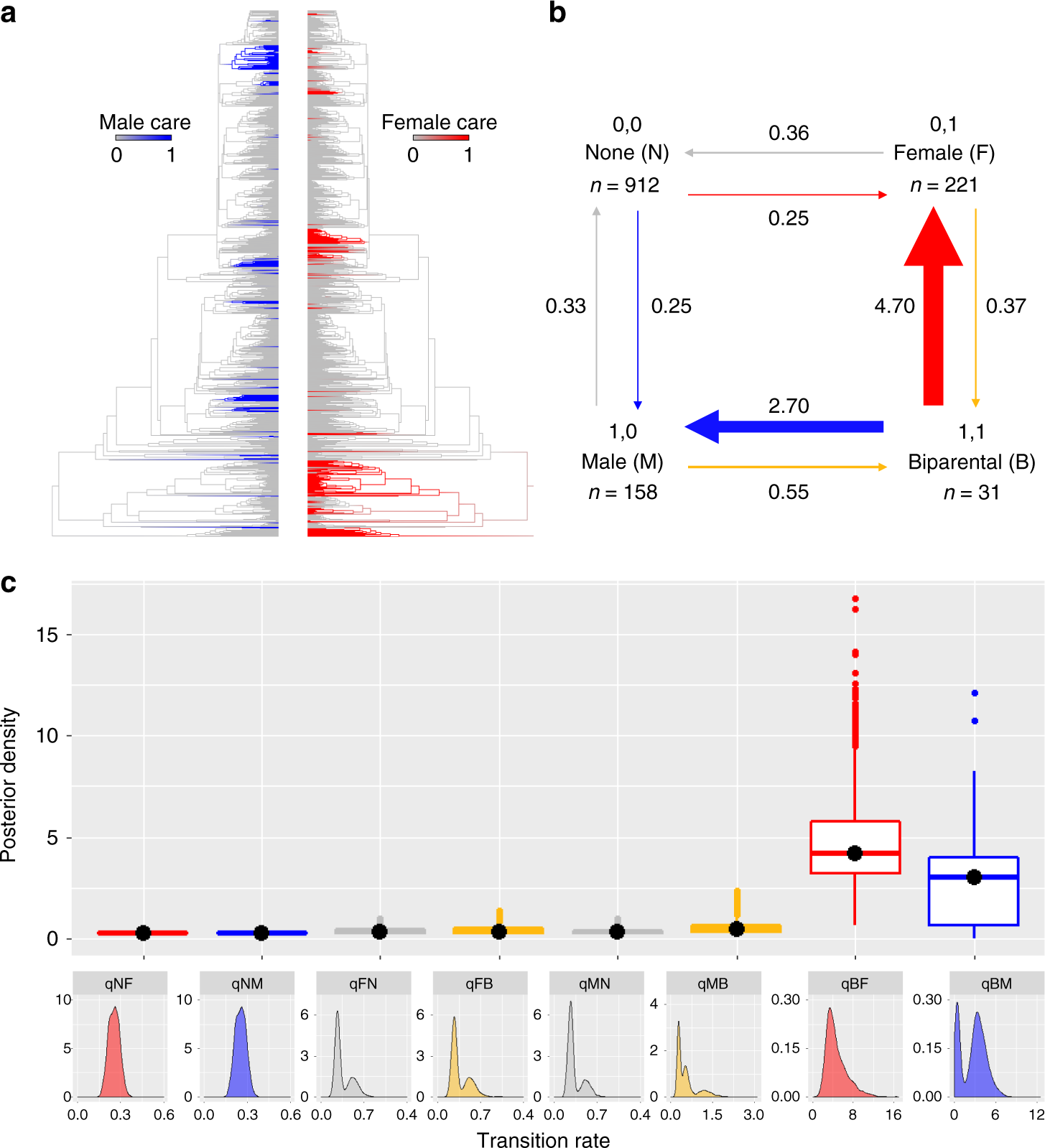
Fig. 4. 兩性撫育行為演化分速率分佈。b, c. 撫育之性別之轉變機率。雌性撫育 (F)。雄性撫育 (M)。兩性共育 (B)。不撫育 (N)。( Furness et al. 2019)
本研究的多樣性尺度大,結果所呈現的趨勢仍很粗略。目前的結論應視為一種大方向的演化趨勢,供後續更詳細的研究進行驗證,並可望在新證據出現後修正此假說。其中關於不同類別的撫育行為消失的速率之預測,可能擴大至昆蟲或魚類等其他演化支系存在。而對於生殖投資與適應性衝突在各支系間的差異,仍須引援其他更細緻的生態模型進行解釋。
References
Furness et al. 2019. Nat. Commun. 10:1-12.
Supplementary information
https://static-content.springer.com/esm/art%3A10.1038%2Fs41467-019-12608-5/MediaObjects/41467_2019_12608_MOESM1_ESM.pdf
Ecology and Evolution
Davies et al. 2014. An introduction to behavioral ecology. Cha8.
Smith 1977 Nature 265: 535-536.
沒有留言:
張貼留言